- Record: found
- Abstract: found
- Article: found
Higher percentage of horse serum in culture media blocks attachment of PC12 cells
letter
16 October 2019
There is no author summary for this article yet. Authors can add summaries to their articles on ScienceOpen to make them more accessible to a non-specialist audience.
Abstract
The rat pheochromocytoma cell line, PC12, has become a major model system to study
many aspects of neuronal activity since its development and characterization in the
1970s [1]. One of its most useful properties is that, following stimulation by nerve
growth factor (NGF), these cells extend processes or neurites in a differentiation
program that is reversible upon NGF removal. The NGF-mediated differentiation includes
the formation of functional synapses as well as electrical excitability [2,3]. Because
of these remarkable properties, PC12 cells have been an important tool to uncover
the mechanisms of neuronal differentiation as well as the NGF signaling pathway. PC12
cells have also been used extensively to study exocytosis, especially of catecholamines
(reviewed in [4]).
According to their initial characterization [1], PC12 cells are round or polygonal,
loosely adherent, and tend to grow in clumps. We report here that their cellular morphology
and adherence is affected greatly by the percentage of horse serum contained in their
culture medium. While the original growth conditions in Greene and Tischler specify
the addition of 10% horse serum and 5% fetal calf serum (FCS) in the growth medium,
which is reinforced by recommendations of the culture repository, ATCC, not all researchers
use the same amount of horse serum in their PC12 cultures. Although 10% horse serum
is most common, amounts ranging between as low as 5% [5] and as high as 15% [6] have
been used. Moreover, reasons for utilizing a specific concentration of horse serum
are routinely not provided and the literature also lacks sufficient explanation as
to its effects. We initially observed, after changing our source of horse serum, that
a larger percentage of cells were more rounded and seemingly loosely attached several
days after plating, which led us to further analyze the effects of horse serum on
the growth of PC12 in culture.
PC12 cells were grown in DMEM media supplemented with increasing percentages of horse
serum (0, 2.5, 5, 10 and 15%). Two different sources of horse serum were used: Donor
Equine Serum from either Corning or HyClone (GE Healthcare Life Sciences). However,
because the focus here is the percentage of horse serum and not variations between
manufacturers (or lots), these two sources are referred to herein simply as horse
serum 1 and 2, in no particular order with respect to the manufacturers named above.
The percentage of cells that were loosely attached (rounded) was determined for each
culture condition, as described in Figure 1. At 24 h post-plating (Figure 1A), as
horse serum percent increased, there was a statistically significant, dose-dependent
increase in the percentage of cells that were loosely attached (p < 0.001 for all
comparisons, except 0 vs 2.5% horse serum, for which p = 0.103). With 10 and 15% horse
serum, the majority of cells were loosely attached at the 24-h time point. At 48 h
post-plating (Figure 1B), with horse serum between 0 and 10%, the majority of cells
were polygonal and attached, with 10% horse serum showing considerably more cells
that were loosely attached than the lower percentages (p < 0.001 in comparison with
0, 2.5 and 5% horse serum). At 15% horse serum, the majority of the cells were loosely
attached at 48 h post-plating (p < 0.001 for all comparisons). These results indicate
that higher percentages of horse serum in the culture media prevent attachment and
spreading of PC12 cells toward a polygonal morphology and lead to more loosely attached
(rounded) cells.
Figure 1.
Percent of loosely attached (rounded) PC12 cells.
PC12 cells were plated in standard culture dishes in media containing the indicated
combination of horse serum and fetal calf serum. Images captured using a Zeiss Primovert
inverted microscope (at 40× magnification) equipped with an Excelis high-definition
digital camera after (A) 24 h and (B) 48 h were analyzed blindly, counting the percentage
of cells loosely attached, as judged by a halo around the cells in the image and a
lack of extensions from the cell body. Cell counts from three images for each combination
were averaged. For statistical analyses, ANOVA with Tukey HSD post-hoc tests were
performed. (Note: In both (A & B), there was no statistically significant difference
between horse serums 1 and 2 at any of the horse serum percentages). (C) A representative
image showing loosely attached cells with halos and attached (polygonal) cells.
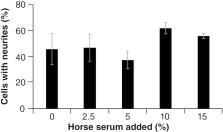
To determine whether the horse serum-mediated differences in cell morphology had a
functional effect on the PC12 cells, neurite outgrowth assays were performed, as described
in Figure 2. Preincubation with the range of horse serum percentages did not show
a statistically significant difference in neurite outgrowth. Although there was substantial
neurite outgrowth following preincubation with all percentages of horse serum (including
0%), there was more variability when using lower percentages (i.e., 0, 2.5 and 5%),
and while the higher percentages (i.e., 10 and 15%) did seem to foster slightly more
neurite outgrowth, there was at best marginal significance, with the comparison of
5 versus 10% horse serum preincubation coming the closest to achieving statistical
significance (p = 0.078). Thus, while variations in horse serum percentage led to
differences in morphology of the PC12 cells prior to NGF-mediated differentiation,
they do not seem to overtly change the ability of these cells to differentiate in
response to NGF.
Figure 2.
Neurite outgrowth assay.
Wells in six-well plates were incubated with 0.5 ml of a 20 μg/ml solution of laminin
(Sigma) and allowed to dry. DMEM media containing the indicated percentages of horse
serum along with 5% fetal calf serum were added to the wells (3 wells for each horse
serum percentage). PC12 cells (1 × 104 cells) were added to each well and allowed
to attach for 48 h. Media was replaced with serum-free DMEM, supplemented with 100 ng/ml
of NGF (Becton Dickenson), and the cells were allowed to incubate for another 48 h.
Five images from each well were captured using a Zeiss Primovert inverted microscope
(at 100× magnification). Cells from each image were counted, both total cells and
those with neurites, defined as neuron-like extensions of the cell membrane that were
equal to or longer than one diameter of the cell. For statistical analyses, ANOVA
with Tukey HSD post-hoc tests were performed.
Overall, the results contained herein suggest that the choice of horse serum percentage
used by researchers is mostly a matter of choice, reflecting their culture preferences.
As the most common percentages of horse serum used seem to be 10 and 15%, which led
to fairly similar performance in neurite outgrowth, the more loosely attached (rounded)
cells seen with 15% horse serum may be preferred by researchers that do not use trypsin
to subculture PC12 cells, allowing them to grow more as a suspension culture. Of note,
our initial observation of changed culture characteristics upon switching horse serum
sources, which caused us to consider discarding the horse serum, can be simply remedied
by altering horse serum concentration.
One question perhaps worthy of further investigation is the effect of long-term culturing
of PC12 cells in medium either devoid of horse serum or containing lower percentages.
The more attached polygonal cells observed with low horse serum percentages (short
term) in this report may be associated with partial differentiation of these cells
toward a non-neuronal cell type that can be reversed by removal of serum and addition
of NGF. It is currently unclear what role the horse serum plays in PC12 cell culture,
but given that PC12s are progenitor cells and can differentiate along both chromaffin
and sympathetic neuronal lineages [1], horse serum may preserve the progenitor status
of these cells. Along these lines, horse serum has also been used to maintain hematopoietic
progenitor cells in culture, keeping them in a less differentiated state [7,8], and
has been seen to block the differentiation of hepatic progenitor cells [9]. Thus,
future studies could further clarify the effect of horse serum on the differentiation
state of PC12 cells.
The discrepancies in PC12 cell attachment seen with different horse serum sources
are likely due to differences in the levels of some factor(s) in these sera. Thus,
another potential avenue of future studies is to determine which components in horse
serum mediate the effect seen on PC12 attachment and are responsible for the variances
observed. Unlike human serum, for which the composition has been characterized [10],
sera used in tissue culture are usually minimally defined and the proteins, lipids,
and/or growth factors and hormones contained in them may vary among sources for many
reasons [11,12]. Ditz et al. were able to determine that high levels of fatty acid
products of phospholipase A2, notably eicosanoids, present in some lots of horse serum
inhibited self-renewal of multipotent murine hematopoietic progenitor cells [8]. While
it is possible that these compounds may similarly affect PC12 cells, identification
of the actual components of horse serum that inhibit attachment (and preserve progenitor
status) is needed, which would allow elucidation of the underlying cellular pathways
and mechanisms as well.
Most cited references12
- Record: found
- Abstract: found
- Article: not found
Establishment of a noradrenergic clonal line of rat adrenal pheochromocytoma cells which respond to nerve growth factor.
L A Greene, A Tischler (1976)
- Record: found
- Abstract: found
- Article: not found
Optimization of chemically defined cell culture media--replacing fetal bovine serum in mammalian in vitro methods.
J. van der Valk, D. Brunner, K De Smet … (2010)
- Record: found
- Abstract: found
- Article: not found
Neuroprotective effect of cannabidiol, a non-psychoactive component from Cannabis sativa, on beta-amyloid-induced toxicity in PC12 cells.
Giuseppe Esposito, Rita Santamaria, Massimo La Rosa … (2004)