- Record: found
- Abstract: found
- Article: found
Low-dose quercetin positively regulates mouse healthspan
letter

Read this article at
There is no author summary for this article yet. Authors can add summaries to their articles on ScienceOpen to make them more accessible to a non-specialist audience.
Abstract
Dear Editor,
Aging is the leading risk factor for many chronic diseases, accounting for almost
60% of all deaths worldwide. How to achieve healthy aging, alleviate aging-related
diseases, and extend healthspan has become a main topic of biomedical research (He
et al., 2019). Geroprotective compounds, such as metformin and rapamycin, have been
shown to improve both healthspan and lifespan in mice (Martin-Montalvo et al., 2013;
Bitto et al., 2016), whereas nicotinamide partially improves healthspan in mice (Mitchell
et al., 2018). In addition, senolytics, compounds that eliminate senescent cells,
have been proven to improve physical function and increase lifespan in mice (Xu et
al., 2018). Although none have proven to be clinically reliable in delaying aging
or treating frailty in humans, these compounds have already provoked enthusiasm for
identifying a potential “elixir”. Therefore, the exploration of more geroprotective
compounds, especially natural active compounds, holds great potential for the development
of geriatric medicines.
Quercetin (Que) is a natural bioflavonoid found in fruits and vegetables such as apples
and onions. Que (50 mg/kg) in combination with dasatinib (5 mg/kg) (abbreviated as
D + Q) has been shown to effectively eliminate senescent cells via induction of apoptosis,
thus alleviating senescence-related phenotypes and improving physical function and
lifespan in mice (Zhu et al., 2015; Xu et al., 2018). In addition, Que (10 mg/kg)
in combination with dasatinib (5 mg/kg) has been reported to reduce hepatic steatosis
(Ogrodnik et al., 2017). In each of these in vivo studies, however, Que was used at
high doses ranging from 10 to 50 mg/kg body weight, which raises concerns about dose-dependent
side effects such as headaches and limb tingling (Shoskes et al., 1999). As a selective
tyrosine kinase receptor inhibitor, dasatinib is associated with warnings and precautions
including pulmonary arterial hypertension and low blood cell counts. Therefore, high-dose
Que and extra side effects of dasatinib would hamper potential clinical applications
of Que in geriatric medicines. Through natural products screening using Werner syndrome
(WS) human mesenchymal stem cells (hMSCs), we recently identified Que as a geroprotective
agent that counteracts accelerated and natural aging of hMSCs at a concentration of
as low as 100 nmol/L, which is 100 times lower than the concentration of Que (10 μmol/L)
previously used in combination with dasatinib as senolytic drugs to eliminate senescent
cells in human umbilical vein cells (HUVECs) (Zhu et al., 2015; Geng et al., 2018).
To explore the geroprotective effect of low-dose Que monotherapy in rodents, we evaluated
the in vivo effect of long-term low-dose Que administration under physiological-aging
condition. Que was given to 14-month-old C57BL/6J male mice by weekly oral gavage
at a concentration of 0.125 mg/kg body weight, which is 80–400 times lower than that
of the previously tested D + Q (10–50 mg/kg body weight) regimens (Fig. 1A), with
vehicle (10% PEG400 in PBS)-treated mice as controls (Zhu et al., 2015; Xu et al.,
2018). After eight months of treatment, Que-treated mice showed decreased hair loss
with normal food intake, body weight, blood glucose and bone mineral density (Figs. 1B
and S1A–D). Compared to vehicle-treated mice, mice subjected to Que treatment showed
markedly improved exercise endurance in the RotaRod and treadmill tests, but normal
grip strength by grip strength meter assay (Figs. 1C, 1D, and S1E–G). Accordingly,
the cardiac function of these mice was examined by Doppler tissue imaging. Although
ejection fraction (EF) and fractional shortening (FS) were unaffected, a higher frequency
of the mitral ratio of peak early to late diastolic filling velocity (E/A) within the
normal range was observed in Que-treated mice than in the age-matched controls (Figs. 1E
and S1H). However, the lifespan was not prolonged by low-dose Que treatment observed
up to the age of 31 months (Fig. S1I). Taken together, these data indicate that long-term
low-dose Que administration alone sufficiently improves multiple aspects of healthspan,
but not lifespan, in mice.
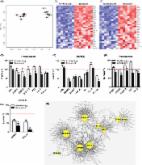
Figure 1
Low-dose quercetin alone improved the healthspan of physiologically aging mice. (A)
Experimental design for drug administration. (B) Hair loss evaluation (n = 11). Data
are shown as the mean ± SEM. *P < 0.05. (C) Hanging endurance on the RotaRod system
(n = 13). Data are shown as the mean ± SEM. *P < 0.05. (D) Frequency of electric shock
on the treadmill over 30 min (n = 12). Data are shown as the mean ± SEM. *P < 0.05.
(E) The ratio of peak velocity of early to late filling of mitral inflow (E/A) (n
= 12). The table shows the number of mice in 3 kinds of E/A ranges, and Que treatment
increased the ratio of normal E/A mice. *P < 0.05. (F) Masson’s trichrome staining
in SKM showed moderate perivascular and interstitial fibrosis (blue areas) (n = 4).
Data are shown as the mean ± SEM. *P < 0.05. Scale bar, 100 μm. SA-β-Gal staining
analysis of SKM, WAT and BAT. Scale bar, 100 μm (n = 4). Data are shown as the mean
± SEM. ***P < 0.001, **P < 0.01, *P < 0.05. Haematoxylin and eosin staining of WAT.
Scale bar, 100 μm (n = 4). Data are shown as the mean ± SEM. *P < 0.05. (G) Global
gene expression profiling in SKM, WAT and BAT (n = 3). Y-Ctrl represents 10-week-old
young male mice, and O-Veh and O-Que represent vehicle (10% PEG400 in PBS)- or low-dose
Que-treated old male mice
To investigate how Que improved healthspan in mice, we collected 11 different kinds
of tissues from 10-week young male mice (Y-Ctrl) and vehicle (O-Veh)- and low-dose
Que-treated 22-month old male mice (O-Que). No significant difference was observed
in organ weights between O-Veh and O-Que (Fig. S2A). Given that exercise endurance
and diastolic function were improved by Que, we particularly examined the changes
in skeletal muscles (SKM), white adipose tissues (WAT), brown adipose tissues (BAT)
and hearts. Upon Que treatment, the arrangement of muscle fibers became more regular
and compact with less fibrosis and senescence (Figs. 1F and S2B). In WAT, the increases
in adipocyte size and senescence-associated β-galactosidase (SA-β-Gal)-positive area
during aging were both alleviated upon Que treatment (Fig. 1F). In BAT, although adipocyte
size was unaffected, there was a decreasing trend of the SA-β-Gal-positive area upon
Que treatment (Figs. 1F and S2B). By comparison, we did not observe any significant
differences in mouse hearts by histological analysis and SA-β-Gal staining (Fig. S2B).
Therefore, these data suggest that long-term low-dose Que administration may delay
aging of SKM, WAT, and BAT in mice.
To further explore the molecular mechanisms of the beneficial effects of Que, we performed
whole-transcriptome RNA sequencing (RNA-seq) of SKM, WAT, and BAT from Y-Ctrl, O-Veh,
and O-Que mice. Global gene expression profiling revealed that most protein coding
genes were unaffected after long-term low-dose Que administration (Fig. 1G). Accordingly,
we inferred that low-dose Que might exert its senostatic effect by regulating the
expression of non-protein-coding RNAs.
We previously observed that Que alleviates hMSC senescence in part through the restoration
of heterochromatin architecture in prematurely and physiologically aging hMSCs (Geng
et al., 2018). Constitutive heterochromatins are predominantly comprised of repetitive
elements (REs), including retrotransposable elements (RTEs). The expression of RTEs
is repressed via epigenetic regulation under normal conditions but is elevated during
physiological aging, eliciting active transposition (De Cecco et al., 2013). Accordingly,
mobilization of RTEs is likely to be a key contributor to tissue aging and cell degeneration
(De Cecco et al., 2013). To investigate whether low-dose Que treatment antagonized
the activation of RTEs, we examined the expression levels of various RTEs, including
long terminal repeats such as LTR10C, LTR2C, LTR35A, and LTR3B, and non-long terminal
repeats including long interspersed nuclear elements 1 (L1, also known as LINE-1)
and short interspersed nuclear elements (SINEs) such as Alu in WS hMSCs after continuous
Que treatment at the concentration of 100 nmol/L. Que treatment silenced the transcription
of various RTEs in WS hMSCs, consistent with the rejuvenated cellular phenotypes (Fig. 2A).
To test whether Que treatment may also repress activation of RTEs in a mouse in vivo
model, we compared the transcriptional levels of RTEs such as L1, SINE B1, LTR41,
LTR42 and MLV5 in multiple tissues of Y-Ctrl, O-Veh, and O-Que mice. Consistently,
most RTEs were transcriptionally upregulated in the SKM and BAT of old mice compared
to those of young mice and were repressed by Que treatment (Fig. 2B). Similar to the
tendency in the BAT, RTEs in WAT from Que-treated mice were also slightly decreased
(Fig. 2B). In line with enhanced L1 transcripts, there was an increased expression
level of L1 open reading frame 1 protein (ORF1p) in SKM and BAT of aged mice, which
could be reversed by long-term low-dose Que administration (Fig. 2C–E). These data
indicate that Que represses RTE activation in senescent hMSCs and multiple aged mouse
tissues.
Figure 2
Activation of retrotransposable elements (RTEs) was repressed in Que-treated WS hMSCs
and certain mouse tissues. (A) RT-qPCR analysis of RTEs in vehicle- and Que-treated
WS hMSCs (passage 7) (n = 3). P values between vehicle and Que are shown on the right.
(B) RT-qPCR analysis of RTEs in SKM, WAT and BAT of young male mice, old male mice
treated with vehicle and Que (n = 4). P values between O-Veh and O-Que are shown on
the right, P ≤ 0.05 were labeled in red. (C) Immunostaining of ORF1p in SKM of young
male mice, old male mice treated with vehicle and Que (n = 3). Scale bar, 50 μm. Data
are shown as the mean ± SEM (cell number ≥ 100). **P < 0.01, *P < 0.05. (D) Immunostaining
of ORF1p in the BAT of young male mice, old male mice treated with vehicle and Que
(n = 3). Scale bar, 7.5 μm. Data are shown as the mean ± SEM (cell number ≥ 100).
***P < 0.001, **P < 0.01. (E) Immunoblotting of ORF1p, RelA, p-TBK1, p-IRF3 and P21
in the SKM and BAT of young male mice (n = 3), old male mice treated with vehicle
(n = 5) and Que (n = 5). ***P < 0.001, **P < 0.01, *P < 0.05, ns, not significant.
(F) Immunostaining of RelA in the SKM of young male mice, old male mice treated with
vehicle and Que (n = 3). Scale bar, 10 μm. Data are shown as the mean ± SEM (cell
number ≥ 100). **P < 0.01, *P < 0.05. (G) Immunostaining of RelA in the BAT of young
male mice, old male mice treated with vehicle and Que (n = 3). Scale bar, 7.5 μm.
Data are shown as the mean ± SEM (cell number ≥ 100). **P < 0.01, *P < 0.05. Y-Ctrl
represents 10-week-old young male mice, and O-Veh and O-Que represent vehicle (10%
PEG400 in PBS)- or low-dose Que-treated old male mice. (H) A proposed model illustrating
the senostatic effects of Que
In senescent cells, the activation of RTEs (such as L1) leads to genome instability
and accumulation of cytosolic DNA that further binds to cytosolic sensor cGAS and
activates TBK1 and IRF3, which subsequently promote senescence-associated secretory
phenotype (SASP) (Takahashi et al., 2018; De Cecco et al., 2019). In addition, NF-κB/RelA
in cGAS-STING-mediated NF-κB pathway acts with IRF3 and other transcription factors
to induce the expression of inflammatory cytokines such as IL-6, the most prominent
SASP cytokine (Chen et al., 2016). Notably, both p-TBK1 and p-IRF3 were increased
in old mouse tissues compared to the young ones and were repressed upon Que treatment
(Fig. 2E), indicating the effect of Que on inhibiting cGAS-STING pathway (Kato et
al., 2017). Similarly, RelA (p65) was upregulated in aged mouse tissues and repressed
upon Que treatment (Fig. 2E–G). Consistently, the inflammatory cytokine IL-6 was increased
in old mice compared to young mice and Que antagonized the increase of IL-6 in both
WS-hMSCs and old mouse SKM and BAT (Fig. 2A and 2B). Thus, our data suggest that Que
may block SASP through the axis of heterochromatin-RTEs (L1)-innate immune response
pathway (Fig. 2H).
In this study, we reported for the first time a geroprotective effect of low-dose
quercetin alone that improved the healthspan of aged C57BL/6J male mice. Que-treated
mice showed less hair loss, greater athletic endurance, enhanced diastolic function,
and less muscle fibrosis, as well as alleviated cellular senescence in multiple tissues.
Interestingly, these changes appear to be rarely associated with transcriptional alterations
of protein-coding genes but are linked to heterochromatin stabilization and RTE silencing.
Que treatment prevented L1 from hyperactivation, thereby inhibiting SASP. In contrast
to the reported senolytic effect of high-dose D+Q (Xu et al., 2018), where Que exerts
geroprotective effects via the induction of apoptosis of senescent cells, low-dose
Que (0.125 mg/kg body weight) alone was sufficient to exert senostatic effects in
mice by affecting heterochromatin stability through repression of RTEs activity in
this study. In a translational context, low-dose Que monotherapy may be helpful to
minimize the dose-dependent side effects compared to high-dose administration and
avoid drug interference when used in combination, probably representing a potential
therapeutic option for future clinical application (Shoskes et al., 1999). Of note,
here we reported the geroprotective effect of Que in male mice and its effect remains
to be studied in female mice.
The possible mechanism of low-dose Que in mice may be associated with its function
as a heterochromatin stabilizer and its direct inhibitory activity against reverse
transcriptase (Ono et al., 1990; Geng et al., 2018). Loss of heterochromatin architecture
and genomic instability are two hallmarks of aging (Zhang et al., 2015). In advanced
age, the expression of RTEs is often increased, which may in turn contribute to genomic
instability and aging-associated cellular defects (De Cecco et al., 2013). Activation
of L1 has been implicated in a variety of age-related disorders, including cancer
and neurodegenerative diseases. The activation of L1 (and possibly other RTEs in mice)
promotes the expression inflammatory factors, a feature of cellular senescence (De
Cecco et al., 2019; Simon et al., 2019). Recently, it has been reported that nucleoside
reverse-transcriptase inhibitors (NRTIs), such as lamivudine, stavudine, inhibit L1
retrotransposition and thus improve the healthspan and/or lifespan of SIRT6-knockout
and physiologically aged mice (De Cecco et al., 2019; Simon et al., 2019). Que has
been proven as a potential inhibitor of reverse transcriptase from Rauscher murine
leukemia virus (RLV) and human immunodeficiency virus (HIV) by enzyme kinetic analysis,
whereas its reverse transcriptase inhibition activity against RTEs in hMSCs and rodents
has not been reported (Ono et al., 1990). Our data provide important evidence supporting
the role of low-dose Que in safeguarding genomic stability (i.e. inhibition of retrotransposition),
which at least in part contributes to its geroprotective activity in rodents.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary material 1 (PDF 438 kb)
Related collections
Most cited references9
- Record: found
- Abstract: found
- Article: found
The Achilles’ heel of senescent cells: from transcriptome to senolytic drugs
Yi-Yi Zhu, Tamara Tchkonia, Tamar Pirtskhalava … (2015)
- Record: found
- Abstract: not found
- Article: not found
LINE1 Derepression in Aged Wild-Type and SIRT6-Deficient Mice Drives Inflammation
Matthew Simon, Michael Van Meter, Julia Ablaeva … (2019)

- Record: found
- Abstract: found
- Article: found