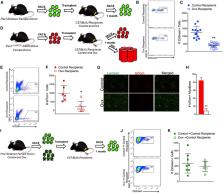
A key determinant of geriatric frailty is sarcopenia, the age-associated loss of skeletal muscle mass and strength 1,2 . Although the etiology of sarcopenia is unknown, the correlation between the loss of satellite cell activity and impaired regenerative capacity in aged muscle has led to the hypothesis that diminished activity of satellite cells, or skeletal muscle stem cells, with age is a cause of sarcopenia 3,4 . We tested this hypothesis using a mouse model to deplete young adult muscle of satellite cells to a level sufficient to impair regeneration throughout the life of the animal. A detailed analysis of multiple muscles in sedentary male mice revealed that, despite reduced regenerative capacity, the life-long reduction of satellite cells did not accelerate nor exacerbate sarcopenia. These data argue against satellite cell contribution to the maintenance of muscle size or fiber type composition during aging; however, an increase in extracellular matrix suggests that loss of satellite cells may contribute to fibrosis with age. Recent estimates indicate that up to one-third of the elderly suffer from frailty, characterized by a common set of symptoms including loss of muscle strength, increased fatigability, modest levels of physical activity and decreased body weight 1 . The close relationship between frailty and the musculoskeletal system suggests sarcopenia is a critical factor contributing to the emergence of geriatric frailty, thus limiting the ability to perform activities of daily living and significantly increasing the risk of falling 5,6 . Numerous studies in humans and rodents report a strong correlation between the loss and dysfunction of satellite cells and sarcopenia 3,4 . Motivated by the idea that the restoration of satellite cell activity will provide a therapeutic basis for treating sarcopenia, a great deal of effort has gone into defining the environmental and cellular changes underlying the loss in satellite cell activity with age 7-18 . Despite the correlation between declining satellite cell-dependent regenerative capacity and age, no studies to date have tested this relationship to determine if the loss of satellite cell activity causes sarcopenia. We recently developed a genetic mouse model that allows for the specific, inducible depletion of satellite cells in adult skeletal muscle 19-21 . The Pax7CreER/+; Rosa26DTA/+ strain, designated Pax7CreER-DTA, was generated by crossing Pax7CreER/CreER and Rosa26DTA/DTA strains. Treatment of the Pax7CreER-DTA mouse with tamoxifen activates Cre recombinase only in satellite cells driven by the Pax7 promoter, which activates the diphtheria toxin A gene, killing satellite cells 21 . We took advantage of this mouse model to directly test the hypothesis that loss of satellite cells, which underlies the well-documented impairment in muscle regenerative capacity 21-24 , results in muscle wasting with advancing age. If there is a causal relationship between the loss of satellite cell activity and sarcopenia, then we would predict accelerated and exacerbated sarcopenia in muscle with a significantly reduced complement of satellite cells. We administered vehicle or tamoxifen by IP injection to adult (4 months of age) male Pax7CreER -DTA mice for five consecutive days to effectively deplete satellite cells and then allowed the mice to age. We analyzed a subset of mice after approximately one year, at 16–18 months of age (middle age, MA), and showed that satellite cell numbers did not recover over time. Consistent with previous studies 21,23 , in muscles which remained significantly satellite cell-depleted (>85%), muscle regeneration following BaCl2 injection was severely impaired (Fig. 1a). No loss of muscle mass was apparent in any hind limb muscle of vehicle- or tamoxifen-treated middle aged mice except in the soleus muscle (MA, Fig.1b); however, significant atrophy was apparent by 24 months in both vehicle- and tamoxifen-treated mice (Aged, Fig. 1b). Decrements in hind limb muscle mass in the aged mice met criteria for sarcopenia in humans 25 ; that is, appendicular muscle mass was two standard deviations below the young group. Thus, loss of satellite cell-dependent regenerative capacity throughout adulthood does not accelerate sarcopenia in aging mice. We next determined if in aged mice, features of sarcopenia were exacerbated due to a lifetime reduction of satellite cells. Analysis of satellite cell abundance in vehicle-treated 5 month old (1 month following injection, young, Fig. 2a) and 24 month old (20 months following injection, aged, Fig. 2a) mice showed a substantial age-associated reduction in satellite cells (Fig. 2b). Tamoxifen administration resulted in >94% reduction in satellite cells in multiple hind limb muscles 1 month following injection, with little recovery in satellite cells per myofiber occurring even after 20 months (Fig. 2b); on average, satellite cells remained 83% depleted in aged mice (range: 64-87%), well below the loss normally associated with aging. Despite the reduction in satellite cell abundance to levels associated with severely impaired muscle regeneration, age-associated atrophy, as indicated both by muscle wet weight (Fig. 1b) and by mean myofiber cross-sectional area (CSA, Fig. 2c), occurred to the same extent in all hind limb muscles from both vehicle-treated and satellite cell-depleted mice. Myofiber CSA analyzed by fiber type (Figs. 3a and b) showed age-related atrophy in all muscles except the soleus, which has a higher proportion of slow-twitch fibers (Fig. 3c). In agreement with the well-characterized atrophy of the largest, fast-twitch glycolytic fibers with age 26 , type 2b fibers showed the greatest degree of atrophy that was unaffected by satellite cell abundance (Fig. 3c). Moreover, fiber size distribution across hind limb muscles showed the characteristic leftward shift due to overall higher abundance of smaller fibers with age in both treatment groups (Supplemental Fig. 1). Satellite cell depletion also had no significant effect on fiber type distribution (Supplemental Fig. 2); at 24 months, only the plantaris and soleus muscles showed an age-dependent shift in the relative frequency of different fast-twitch (type 2) fibers that was independent of satellite cell abundance. In addition to atrophy of individual myofibers, sarcopenia is associated with a decline in the number of myofibers and a reduction in single fiber specific force generation (force per unit area) 2 . We measured these characteristics in the plantaris muscle, as it showed little recovery of satellite cells with age and was therefore most severely depleted (see Fig. 2b). We did not observe a loss of myofibers at 24 months in either vehicle- or tamoxifen-treated mice (Fig. 4a). Analyses of isolated myofibers (Fig. 4b) showed that the number of myonuclei per 100 μm of myofiber length was not changed with age (Fig. 4c). We performed myonuclear counts primarily on intermediate-sized myofibers (1000–2500 μm2), that make up greater than 80% of all myofibers, and our results are consistent with a recent study reporting no myonuclear loss with age in fibers in this size range 27 . Moreover, myonuclear number was unaffected by satellite cell depletion (Fig. 4c), suggesting that the maintenance of myonuclei is not linked to satellite cell abundance. This conclusion was further supported by analysis of BrdU-labeled myonuclei (Fig. 4d). We provided mice with BrdU via drinking water for two weeks prior to sacrifice, and quantified fusion of labeled nuclei into myofibers. Although myonuclear addition was very infrequent during the two week labeling (approximately 0.1% of myofibers from vehicle-treated young and aged mice contained a labeled myonucleus), no labeled myonuclei were detected in plantaris muscle following tamoxifen treatment (Fig. 4d). Finally, as shown in Fig. 4e, myofiber specific force was reduced 31% with age, comparable to the force decrement reported in isolated myofibers in adult humans aged 65–85 years, using a similar permeabilized fiber preparation 28 ; however, the decrease in specific force was unaffected by satellite cell abundance. We also functionally tested single fibers from the EDL muscle from 24 month old tamoxifen- and vehicle-treated mice (Supplemental Fig. 3a–c). As with the plantaris muscle, neither specific or absolute force, nor single fiber CSA, were affected by satellite cell depletion in the EDL. Further, overall loss of function with age, as measured by grip strength, was not altered by satellite cell depletion (Supplemental Fig. 3d). The loss of 45–59% raw grip strength in the aged mice did not differ with treatment, and is consistent with loss reported in humans with age 29,30 . Although age-related myofiber atrophy and weakness were not altered by lifelong satellite cell reduction, we did observe a change in the muscle fiber environment. Extracellular matrix (ECM) surrounding myofibers, quantified by Sirius Red staining of collagens, was higher with age and reduced satellite cell content in the plantaris muscle (representative images, Fig. 4f and g, quantified in Fig. 4h). We also assessed ECM accumulation via alpha-wheat germ agglutinin (WGA) staining of glycosaminoglycans, which showed that this component of the ECM was also more abundant with age and satellite cell depletion (representative images Supplemental Fig. 4a and b, quantified in Supplemental Fig. 4c). WGA staining of other hind limb muscles showed excess accumulation of ECM with age that was further exacerbated by a reduction in satellite cell content specifically in the plantaris and TA/EDL muscles (Supplemental Fig. 4c). These observations support our recent findings that satellite cells regulate the myofiber environment by signaling to fibroblasts 19 . While our previous work illustrated an additive effect of satellite cell-depletion and functional overload on ECM deposition in young adult mice 19 , novel findings in the current study show that long term depletion of satellite cells in sedentary mice contributes to dysregulation of the ECM in old age. Moreover, the present work extends previous findings to suggest that satellite cells may limit fibrosis preferentially in fast muscles 31 . In summary, although our understanding of satellite cell function in muscle regeneration continues to be refined, the present study provides convincing evidence that the loss of satellite cell-dependent regenerative capacity neither accelerates nor exacerbates sarcopenia. Moreover, our findings have broader implications for the study of tissue homeostasis by showing that skeletal muscle, much like the pancreas, kidney and liver, apparently employs cellular mechanisms that do not necessarily require stem cell participation for tissue maintenance 32-35 . However, the loss of satellite cells may adversely affect overall muscle quality, potentially contributing to the increase in fibrosis observed in aged skeletal muscle 12 . One limitation of the current findings stems from the sedentary nature of the mice. The satellite cell requirement for muscle maintenance in more physically active mice remains to be determined; however, the sedentary nature of the mice in the current study is reflective of low physical activity levels of older adults in the United States 36 . The findings of the current study have clinical importance as they draw a clear distinction between therapeutic strategies that may effectively treat degenerative myopathies, such as dystrophies 37 and cachexia 38 , versus sarcopenia. While degenerative conditions are expected to benefit from a satellite cell-based therapy, our results support the conclusion of a recent study 39 , that treatment for sarcopenia should more appropriately focus on the myofiber and motor neuron. Online Methods Mouse Model The Pax7CreER-DTA genetic mouse model allows for the specific and inducible depletion of satellite cells upon tamoxifen treatment, through activation of the diphtheria toxin A gene only in Pax7-expressing cells 21 . The Pax7CreER strain was generated by placing an ires-CreERTM-FRT-Neo-FRT cassette into the ClaI site of the Pax7 gene 3′-UTR following the stop codon in exon 9 40 . Mice are on a mixed C57BL/6 – 129 background strain. All animal procedures were conducted in accordance with institutional guidelines approved by the Institutional Animal Care and Use Committee of the University of Kentucky. Mice were housed in a temperature- and humidity-controlled room and maintained on a 14:10 h light: dark cycle with food and water ad libitum. Mice were not housed in a barrier/pathogen-free facility, and were not screened for tumors. Sample size was determined by a power analysis with an expected standard deviation of 0.001, power of 0.8 and alpha of 0.05. Adult (4 months of age), male Pax7CreER-DTA mice were randomized and administered by intraperitoneal (IP) injection either vehicle (15% ethanol in sunflower seed oil) or tamoxifen (2.0 mg · d−1) for five consecutive days, two hours prior to lights out. Following one month (5 month old, young), 12 months (16–18 month old, middle aged (MA)) or 20 months (24 month old, aged), vehicle and tamoxifen-treated mice were sacrificed (n=4–8 mice/group). Aged vehicle mice had a median lifespan of 23.9 months, and a maximum lifespan of 24.6 months; aged tamoxifen mice had a median lifespan of 22.6 months and a maximum lifespan of 24.4 months. A subset of mice was provided with 5-bromo-2′-deoxyuridine (BrdU) at a concentration of 0.8 mg · ml−1 in their drinking water for two weeks prior to sacrifice. BaCl2-induced muscle injury MA mice were anesthetized with isoflurane and the tibialis anterior (TA) injected with either 50 μl of 1.2% BaCl2 solution or sterile PBS. After seven days, TA muscles were collected and processed for histochemistry. Histochemistry/Immunohistochemistry Muscles were mounted at resting length, covered in OCT compound and frozen in liquid nitrogen-cooled isopentane and stored at −80 °C until cryosectioning (7μm). For Pax7 (satellite cells) immunohistochemistry, muscle sections were fixed in 4% paraformaldehyde (PFA) followed by epitope retrieval using sodium citrate (10 mM, pH 6.5) at 92 °C for 20 min. Endogenous peroxidase activity was blocked with 3% hydrogen peroxide in phosphate-buffered saline followed by an additional blocking step with Mouse-on-Mouse Blocking Reagent (Vector Laboratories, Burlingame, CA). Incubation with Pax7 antibody (1:100, Developmental Studies Hybridoma Bank, Iowa City, IA) was followed by incubation with the biotin-conjugated secondary antibody and streptavidin-HRP included within the Tyramide Signal Amplification kit (Cat#T20935, Invitrogen, Carlsbad, CA). Sections were co-stained with DAPI (4′, 6′-diamidino-2-henylindole, 10 nM, Invitrogen), and mounted with Vectashield fluorescent mounting media. For fiber typing, unfixed sections were incubated in antibodies against myosin heavy chain (MyHC) types 1, 2a and 2b (1:100, Cat#BA.D5, SC.71 and BF.F3, Developmental Studies Hybridoma Bank) in addition to dystrophin (1:50, Cat#VPD505, Vector). MyHC type 2x expression was assumed from unstained fibers. Fluorescent-conjugated secondary antibodies against various mouse immunoglobulin subtypes were applied to visualize MyHC expression and dystrophin. Sections were post-fixed in 4% PFA prior to mounting. For extracellular matrix accumulation, muscle sections were fixed in 4% PFA, and then incubated with Texas-red conjugated WGA (wheat germ agglutinin, Cat#W21405, Invitrogen). Basic muscle morphology and regeneration were assessed with hematoxylin and eosin staining, and collagen content was assessed with Sirius Red staining following standard protocols. For BrdU detection, unfixed slides were incubated in an antibody against dystrophin followed by secondary antibody conjugated to Texas-Red (Cat#610-109-121, Rockland Immunochemicals Inc., Gilbertsville, PA). Sections were then fixed in absolute methanol, treated with 2 N HCl to denature DNA and neutralized with 0.1 M borate buffer (BORAX, pH 8.5). BrdU antibody incubation was followed by biotin-conjugated goat anti-mouse secondary antibody and streptavidin-FITC (Cat#SA-5001, Vector). Sections were postfixed in PFA and co-stained with DAPI. Image Quantification Images were captured at 10 and 20x with an upright microscope (AxioImager M1; Zeiss, Göttingen, Germany). Fiber type-specific cross-sectional area was quantified using a newly developed, automated image segmentation algorithm 41,42 , that identifies fiber types by MyHC isoform expression combined with fiber boundary detection using dystrophin immunohistochemistry. All other images were quantified with Zeiss Axiovision rel. software (v4.8). Satellite cell abundance was assessed using Pax7 staining and only those cells that were Pax7+ and DAPI+ were counted. Fibers were classified as BrdU+ with a BrdU+/DAPI+ nucleus within the dystrophin border. WGA and Sirius Red staining were quantified using threshold intensity programs within the imaging software. Investigators were blinded to treatment (vehicle/tamoxifen) but not age during image quantification and analysis. Single Myofiber Contractility Measurement of permeabilized muscle fiber contractility was performed as previously described 43 . Muscles were removed from the animal and immediately placed in ice cold relaxing solution. Bundles of fibers were dissected from whole muscles, placed in skinning solution for 30 min and then in storage solution for 16 h at 4 °C and assayed immediately or stored at −80 °C until use. Individual myofibers (n=8 myofibers/mouse; 3-8 mice/group; see figure legends for specific numbers) were pulled from bundles in relaxing solution and secured at one end to a force transducer (Aurora Scientific, Model 403, Ontario, Canada) and at the other end to a servomotor (Aurora Scientific). The length of the whole fiber was adjusted to obtain a sarcomere length of 2.5μm using laser diffraction techniques. The average fiber cross-sectional area (CSA) was calculated assuming an elliptical cross-section, with diameters obtained at five different positions along the fiber from high-magnification images of the top and the side views. Maximum isometric force (Fo) of the fiber was elicited by immersing the fiber in a high-Ca2+ concentration solution. Specific force (sFo) was calculated by dividing Fo by CSA. Myonuclear Number Plantaris muscles were fixed in situ at resting length in 4% PFA for 48 h. Single myofibers were isolated by 40% NaOH digestion, as previously described 21 . Single myofibers were stained with DAPI and nuclei from 15–25 myofibers per animal (n = 4–8 mice/group) within a given segment were counted by z-stack analysis using the AxioImager M1 microscope. AxioVision software was used to measure myonuclear number per myofiber segment. Grip Strength Mice were held at the base of the tail and were allowed to grab with fore limb paws the horizontal bar of the grip strength apparatus in a pronated manner (Columbus Instruments, Columbus, OH) while being held horizontally. The mice were then pulled back gently until the grip was released and the maximal force achieved by the animal was recorded (in N); hind limbs were kept free from the apparatus and the average of 3 trials was reported. Data are presented as mean grip strength per body weight. All mice were tested by the same person. Statistics All data were analyzed with SigmaPlot v12.0 software (Systat Software, San Jose, CA) via a two-factor ANOVA (vehicle/tamoxifen x young/MA/aged) or (vehicle/tamoxifen x young/aged) or simple two-tailed t-tests for each dependent variable under consideration. Assumptions for statistical analyses were met (i.e. normal distribution, equal variance). If a significant interaction was detected, an appropriate post-hoc analysis was employed to determine the source of the significance. Statistical significance was accepted at P < 0.05. Data are reported as mean ± standard error. Supplementary Material 1