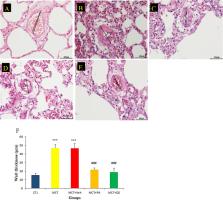
Pulmonary arterial hypertension (PAH) is a vascular disease that is largely restricted to small pulmonary arteries (PAs). PAH occurs in rare idiopathic and familial forms, but is more commonly part of a syndrome associated with connective tissue diseases, anorexigen use, HIV, or congenital heart disease. Many abnormalities contribute to this syndrome of obstructed, constricted small PAs. This includes abnormalities in the blood content of some neurotransmitters and cytokines, namely increases in serotonin, IL-6, platelet-derived growth factor (PDGF), and endothelin (Stewart et al., 1991; Christman et al., 1992; Steudel et al., 1997; Perros et al., 2008). The media is also characterized by an increased activation of the nuclear factor of activated T cells (NFAT), leading to increased [Ca2+]i-mediated PA smooth muscle cell (PASMC) proliferation and decreased mitochondrial-dependent apoptosis (Bonnet et al., 2006, 2007b). Finally, the adventitia is infiltrated with inflammatory cells and exhibits metalloprotease activation (Humbert et al., 2004). Despite recent therapeutic advances such as endothelin-1 receptor blockers (e.g., Bosentan; Dupuis and Hoeper, 2008), type 5 phosphodiesterase inhibitors (e.g., sildenafil; Li et al., 2007), or PDGF receptor blockers (e.g., imatinib; Ghofrani et al., 2005), mortality rates remain high (Archer and Rich, 2000). Moreover, the fact that the PAH phenotype is preserved in cultured PASMCs isolated from PAH patients suggests that the PAH phenotype is sustained independently of the circulating growth factors or agonists but requires genetic remodeling processes (Yildiz, 2009; Dumas de la Roque et al., 2010). Over the past 10 yr, genetic research on PAH has led to the discovery of mutations in the BMPR2 (bone morphogenetic receptor-2) gene in at least 50% of familial PAH patients, and its down-regulation is recognized as a hallmark of PAH (Tada et al., 2007; Zakrzewicz et al., 2007). Recently, BMPR2 down-regulation in human PASMCs has been linked to the activation of both the tyrosine kinase Src (Wong et al., 2005) and a STAT3/miR-17-92 microRNA (miRNA) secondary to IL-6 exposure, suggesting the implication of miRNAs in the etiology of PAH (Brock et al., 2009). miRNAs are small noncoding RNAs (21–23 nt) that are now known to be important regulators of gene expression. They form imperfect RNA–RNA duplexes and use their seed region to interact with messenger RNAs (mRNAs), mainly in the 3′ untranslated region (UTR; Khan et al., 2009). This interaction leads to a negative posttranscriptional regulation of the relevant mRNAs. Recently, misexpression of miRNAs has been implicated in many cardiovascular diseases, including pulmonary hypertension (Latronico and Condorelli, 2009; Mishra et al., 2009; Zhang, 2009; Caruso et al., 2010), but their molecular role in these pathologies has not been uncovered yet. RESULTS miR-204 is aberrantly expressed in human PAH-PASMCs To determine whether miRNAs are aberrantly expressed in human PAH, PASMCs were isolated from distal PAs of two nonfamilial PAH patients (two idiopathic PAH [iPAH] patients A and B; based on the World Health Organization [WHO] classification) and two control patients (A and B) and cultured as previously described (passage 3 and less; McMurtry et al., 2005). The expression of 377 different miRNAs was measured. Seven miRNAs were aberrantly expressed in PAH-PASMCs compared with control PASMCs (miR-204, -450a, -145, -302b, -27b, -367, and -138; Fig. S1 A). Among them, only the miR-204 level was down-regulated (Fig. S1 A). miR-204 down-regulation between control versus PAH was confirmed by quantitative RT-PCR (qRT-PCR) in PASMCs isolated from three PAH (all from group 1 based on WHO classification patients A–C) compared with control PASMCs isolated from five control patients (A–E). Note that no significant differences in miR-204 expression was found among the control patients and the PAH patients (Fig. S1 B). Therefore, for the rest of the study, all five control PASMC and the three PAH-PASMC cell lines were used for every cell-based experiment. Interestingly, in retinal epithelial cells and several cancer cells, miR-204 down-regulation has been associated with enhanced cell proliferation and membrane potential depolarization (Lee et al., 2010; Wang et al., 2010), which are both aspects also seen in PAH-PASMCs (Bonnet et al., 2006, 2007b). We recently showed in several cancer cells and PAH-PASMCs (Bonnet et al., 2007a,b) that this pro-proliferative phenotype was associated in part with the activation of the Src–STAT3 (accounting for BMPR2 down-regulation; Wong et al., 2005) and NFAT pathways (Bonnet et al., 2007b). This suggests a putative link between miR-204 down-regulation, NFAT activation, and cell proliferation. Thus, miR-204 is likely implicated in PAH, and a role in the sustainability of the PAH-PASMC pro-proliferative and antiapoptotic phenotype is possible. Therefore, the current study will be focused on the role of miR-204 in the etiology of PAH. Interestingly, using in silico and microarray gene expression analyses, we observed that among the 461 predicted targets of miR-204 (TargetScan 5.1), only 165 were increased by artificial miR-204 inhibition in control human PASMCs (n = 2 patients; Fig. S1 C). In accordance with the pro-proliferative and antiapoptotic phenotypes seen in PAH, several Src–STAT3- and NFAT-related genes were identified (Fig. S1 C). miR-204 expression is decreased in human PAH and correlates with PAH severity To investigate the expression pattern of miR-204 in normal and pulmonary hypertensive lungs, we examined miR-204 expression levels in (a) lung biopsies from 8 individuals with nonfamilial PAH compared with biopsies from 8 individuals without pulmonary hypertension, (b) lungs from 6 mice with hypoxia-induced pulmonary hypertension compared with 5 control littermates, and (c) lungs from 5 rats with monocrotaline (MCT)-induced pulmonary hypertension compared with 10 control littermates (Fig. 1 A). We found decreased levels of miR-204 in human and rodent pulmonary hypertensive lung tissues compared with normotensive lung samples. To characterize whether down-regulated miR-204 levels were specific to the lung in rats with pulmonary hypertension, we compared organ-specific levels of miR-204 between normal and pulmonary hypertensive rats (Fig. 1 B). Even if we were able to detect minimal amounts of miR-204 in most organs, miR-204 levels were only down-regulated in the lung and PAs but not in the aorta, liver, heart, and kidney in rats 3 wk after MCT injection (pulmonary hypertensive rats) compared with non–pulmonary hypertensive rats (Fig. 1 B). Figure 1. Correlation between miR-204 expression and PAH severity. (A) miR-204 is decreased in human, mouse, and rat PAH lungs. qRT-PCR analysis of miR-204 expression in human lungs with PAH (n = 8), mouse lungs with hypoxia-induced pulmonary hypertension (n = 6), and rat lungs with MCT-induced pulmonary hypertension (n = 5) compared with human (n = 8), mouse (n = 10) and rat (n = 5) control (Ctrl) lungs. (B) miR-204 is mainly expressed in the distal PAs. qRT-PCR analysis of miR-204 expression in several rat organs with MCT-induced pulmonary hypertension (n = 5) compared with control rats (n = 5). (C) miR-204 down-regulation correlates with PAH severity. qRT-PCR analysis of miR-204 expression in the lungs from healthy subjects (n = 8) and from patients with varying severity of PAH (n = 3), in mouse lungs with varying severity of hypoxia-induced pulmonary hypertension (n = 3), and in rat lungs with varying severity of MCT-induced pulmonary hypertension (n = 3) compared with control animals (n = 5 for both rats and mice; n = 3 experiments per patient or per animal for each pulmonary vascular resistance [PVR] or mean pulmonary arterial pressure [PAP] listed, and significance is compared with control group). In all experiments, the level of miR-204 is relative to the control RNA U6. Data are expressed as means ± SEM (*, P 25 mmHg). Age- and sex-matched control PASMCs (three males A, B, and C 45, 21, and 64 yr old; and two females D and E 17 and 35 yr old), and PAECs were purchased from Cell Application USA. PASMCs were grown in high-glucose DME supplemented with 10% FBS (Invitrogen) and 1% antibiotic/antimitotic (Invitrogen; Bonnet et al., 2007a). STAT3 and SHP2 were inhibited by a specific siRNA (20 nM for 48 h; Applied Biosystems) as previously described (Bonnet et al., 2007a). NFAT was inhibited by 4 µM VIVIT as previously described (Bonnet et al., 2007b). Control PASMCs were exposed to 30 ng/ml PDGF, 10 nM endothelin-1, 200 nM angiotensin II, or 100 ng/ml TNF (all from EMB Canada). The Src inhibitor PP2 effects were compared with its negative control inhibitor PP3 (4-amino-7-phenylpyrazol [3,4-d] pyrimidine; 10 µM for 48 h). The Ca phosphate transfection method was used to transfect miRIDAN miR-204 mimics (200 nM for 48 h) or miR-204 antagomir (hairpin inhibitor at 200 nM for 48 h). For each experiment, we used a proper control (mimics or hairpin inhibitor negative control #1 from Thermo Fisher Scientific). Dose response, transfection efficiency, and siRNA efficiencies are presented in Fig. S8 C. TaqMan low density arrays (TLDAs). TLDA was performed in four patients, two for each condition, and according to the manufacturer’s protocol (Applied Biosystems). Each sample was analyzed in duplicate. Raw CTs were then normalized using two different normalization procedures, one normalizing relative to U6 small nuclear RNA and the other one relative to the median CT. An empirical Bayesian method within the package limma in Bioconductor was used to identify the significantly modulated miRNAs. miRNAs were required to be significantly modulated for both normalizations. TLDA data have been deposited in GEO DataSets under accession no. GSE21284. DNA microarrays. DNA microarray experiments were performed using the Whole Human Genome microarray kit (Agilent Technologies). The arrays were scanned using a dual-laser DNA microarray scanner (Agilent Technologies), and the data were extracted from images using the Feature Extraction software. For the control versus PAH patient comparison, RNAs extracted from two control patients were hybridized on Cy3, and RNAs extracted from two PAH patients were hybridized on Cy5. For the miR-204 inhibition experiment, RNAs extracted from control PASMCs treated for 48 h with 200 nM miR-204 antagomir (Thermo Fisher Scientific) were hybridized with Cy3, whereas PASMCs treated with antagomir negative were hybridized on Cy5. Data were background subtracted and normalized within the array using the LOESS normalization before significant modulation assessment using the Empirical Bayes method within limma in Bioconductor. Genes listed as targets of miR-204 in TargetScan 5.1 and having a level of expression ≥100 in log2 base and being up-regulated after miR-204 inhibition were considered as miR-204 targets in our model. Microarray data have been deposited in GEO DataSets and are available under accession no. GSE21284. qRT-PCR. To measure miR-204 expression, the mirVana kit (Applied Biosystems) was used to extract total RNA from PAH-PASMCs or control PASMCs. Stem-loop qRT-PCR for mature miRNAs was performed on a real-time PCR system (AB 7900; Applied Biosystems). Regular qRT-PCR was performed as previously described (Bonnet et al., 2007b). ChIP-PCR. In brief, control PASMC asynchronously growing cells were treated with endothelin at 10 nM. Cross-links were generated with 1% formaldehyde, and chromatin was extracted in lysis buffer (50 mM Tris-HCl, pH 8, 10 mM EDTA, 0.2% SDS, and 5 mM Na-butyrate). Chromatin was then sheared by sonication (Bioruptor; Diagenode) on ice to a mean length of 750 bp. After preclearing with a mix of protein A/G–Sepharose beads (4°C for 1 h), 80 µg chromatin was used for immunoprecipitation with appropriate antibodies (10 ml p-Sat3 [Tyr705; 9131; Cell Signaling Technology] and 10 mg normal rabbit IgG [I-1000; Vector Laboratories]) in a total volume of 300 ml. After overnight incubation at 4°C, 25 µl of protein A Dynabeads (Invitrogen) was added and incubated for >1 h. Beads were extensively washed, and immunoprecipitated complexes were eluted in buffer E (50 mM Na bicarbonate and 1% SDS). Cross-links were reversed overnight at 65°C. Samples were treated with proteinase K, and the DNA was extracted using phenol-chloroform. Quantitative real-time PCR was performed using SYBR green I (LightCycler 480; Roche). Enrichment for a specific DNA sequence was calculated using the comparative Ct method. The numbers presented with standard errors are based on two biological repeats (cells/chromatin/immunoprecipitation). Primers used in the PCR reactions (Table S2) were analyzed for specificity, linearity range, and efficiency to accurately evaluate occupancy (percentage of immunoprecipitation/input). Vascular endothelial growth factor (VEGF) primers were used as positive control, whereas OR8J1 primers were used as negative control. Confocal microscopy. NFATc1 and -c2 and STAT3 nuclear translocation assays were performed using antibodies (1:250; Abcam) as previously described (Bonnet et al., 2007b). TMRM, TUNEL, PCNA, and Fluo-3 were measured as previously described (Bonnet et al., 2009; Bonnet et al., 2007b). Transfection and luciferase assay for different 3′ UTR constructions. The 3′ UTRs of each gene of interest were cloned and inserted in the psiCHECK2 plasmid immediately downstream from the stop codon of firefly luciferase. Once ready, cells were transfected with the reporter plasmid with 200 nM of unrelated small RNA duplex (mimic control; Invitrogen), miR-204 mimic (Thermo Fisher Scientific), miR-204 inhibitor (Thermo Fisher Scientific), or with Caenorhabditis elegans miR-67 inhibitor used as control (Thermo Fisher Scientific). Firefly and Renilla luciferase activities were measured consecutively using a dual-luciferase assay 48 h after transfection. The presence of an interaction between miR-204 and target mRNA would reduce the firefly luciferase activity (normalized to Renilla luciferase activity expressed from the psiCHECK2 plasmid). To abrogate miR-204 binding to the SHP2 3′ UTR, point mutations were made into the 3′ UTR sequence that corresponds to the miR-204 binding site in position 2, 4, and 6 from the 5′ end of miR-204, as illustrated in Fig. 4 B. In vivo model rats. Male rats were injected s.c. with a 60-mg/kg MCT solution (Todorovich-Hunter et al., 1988). PAH was assessed by hemodynamic measurements (using Swan-Gan catheters) and echocardiography (using Vevo 2100; VisualSonics), which were performed as previously described (Bonnet et al., 2007b). In vivo, rats with established PAH (measured by Echo-Doppler) were nebulized with miR-204 mimic (mature sequence, 3′-UUCCCUUUGUCAUCCUAUGCCU-5′) or mimic negative (20 µM once a week for 2 wk). Invivofectamine (Invitrogen) was used as transfected agent according to the manufacturer’s instructions. Transfection efficiency and tissue distribution were assessed by qRT-PCR. Tissue distribution was assessed using fluorescent distribution of the commercially available DY547-labeled mimic control (Thermo Fisher Scientific). Chronic hypoxic mice model. Mice were placed for 2–3 wk in normobaric hypoxic chambers maintained with 5.5–liter min−1 flow of hypoxic air (10% O2 and 90% N2). Chambers were opened twice a week for cleaning and replenishment of food and water. Oxygen concentration was continuously monitored with blood gas analyzers. Soda lime was used to lower carbon dioxide concentration. Statistical analysis. Values are expressed as fold change or mean ± SEM. Unpaired Student’s t tests were used for comparisons between two means. For comparisons between more than two means, we used one-way analysis of variance followed by a Dunn’s test. A p-value <0.05 was considered statistically significant (and indicated with asterisks). TLDA and microarray data were analyzed within R (http://www.r-project.org) using the limma package in Bioconductor. Online supplemental material. Fig. S1 shows seven miRNAs that are aberrantly expressed in human PAH-PASMCs compared with control PASMCs. Fig. S2 represents the measurements of miR-204 level in the pulmonary vasculature and buffy coat. Fig. S3 shows that miR-204 down-regulation in PAH-PASMCs promotes the activation of STAT3 and NFAT. Fig. S4 shows that the miR-204 effect is independent of TRPM3 expression. Fig. S5 shows that the miR-204 mimic molecule restores BMPR2 expression in PAH. Fig. S6 shows that a decrease of miR-204 level activates the Src–STAT3 axis and promotes NFAT expression. Fig. S7 shows that miR-204 mimic intratracheal nebulization restores miR-204 expression in distal PAs, which improves the PAAT and decreases right ventricle hypertrophy in the MCT rat model. Fig. S8 shows the validation of miR-204 mimic/antagomir transfection and siRNA effects on PASMCs. Table S1 lists patients providing tissue. Table S2 lists primers used for ChIP–real-time PCR. Online supplemental material is available at http://www.jem.org/cgi/content/full/jem.20101812/DC1.