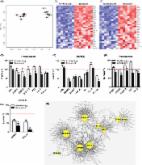
Although recent studies have revealed that heart cells are generated in adult mammals, the frequency and source of new heart cells is unclear. Some studies suggest a high rate of stem cell activity with differentiation of progenitors to cardiomyocytes 1 . Other studies suggest that new cardiomyocytes are born at a very low rate 2,3,4 , and that they may be derived from division of pre-existing cardiomyocytes. Thus, the origin of cardiomyocytes in adult mammals remains unknown. Here we combined two different pulse-chase approaches -- genetic fate-mapping with stable isotope labeling and Multi-isotope Imaging Mass Spectrometry (MIMS). We show that genesis of cardiomyocytes occurs at a low rate by division of pre-existing cardiomyocytes during normal aging, a process that increases by four-fold adjacent to areas of myocardial injury. Cell cycle activity during normal aging and after injury led to polyploidy and multinucleation, but also to new diploid, mononucleated cardiomyocytes. These data reveal pre-existing cardiomyocytes as the dominant source of cardiomyocyte replacement in normal mammalian myocardial homeostasis as well as after myocardial injury. Despite intensive research, fundamental aspects of the mammalian heart’s capacity for self-renewal are actively debated 5,6 . Estimates of cardiomyocyte turnover range from less than 1% per year 2,3,4 to more than 40% per year 7 . Turnover has been reported to either decrease 3 or increase with age 7 , while the source of new cardiomyocytes has been attributed to both division of existing myocytes 8 and to progenitors residing within the heart 9 or in exogenous niches such as bone marrow 10 . Controversy persists regarding the plasticity of the adult heart in part due to methodological challenges associated with studying slowly replenished tissues. Toxicity attributed to radiolabeled thymidine 11 and halogenated nucleotide analogues 12 limits the duration of labeling and may produce direct biological effects. Tissue autofluorescence can reduce the sensitivity and specificity of immunofluorescent methods of detecting cell cycle activity 5,13 , including as cell cycle markers or incorporation of halogenated nucleotide analogues. The challenge of measuring cardiomyocyte turnover is further compounded by the faster rate of turnover of cardiac stromal cells relative to cardiomyocytes 14 . We used Multi-isotope Imaging Mass Spectrometry (MIMS) to study cardiomyocyte turnover and to determine whether new cardiomyocytes are derived from preexisting myocytes or from a progenitor pool (Fig 1a). MIMS uses ion microscopy and mass spectrometry to generate high resolution quantitative mass images and localize stable isotope reporters in domains smaller than one micron cubed 15,16,17 . MIMS generates 14N quantitative mass images by measuring the atomic composition of the sample surface with a lateral resolution of under 50nm and a depth resolution of a few atomic layers. Cardiomyocyte cell borders and intracellular organelles were easily resolved (Fig 1b). Regions of interest can be analyzed at higher resolution, demonstrating cardiomyocyte-specific subcellular ultrastructure, including sarcomeres (Fig 1c, Supplemental Fig 1a). In all subsequent analyses, cardiomyocyte nuclei were identified by their location within sarcomere-containing cells, distinguishing them from adjacent stromal cells. An immense advantage of MIMS is the detection of nonradioactive stable isotope tracers. As an integral part of animate and inanimate matter, they do not alter biochemical reactions and are not harmful to the organism 18 . MIMS localizes stable isotope tracers by simultaneously quantifying multiple masses from each analyzed domain; this enables the generation of a quantitative ratio image of two stable isotopes of the same element 15 . The incorporation of a tracer tagged with the rare stable isotope of nitrogen (15N) is detectable with high precision by an increase in 15N:14N above the natural ratio (0.37%). Nuclear incorporation of 15N-thymidine is evident in cells having divided during a one-week labeling period, as observed in the small intestinal epithelium, which turns over completely in 3–5 days 16 (Fig 1d); in contrast, 15N-thymidine labeled cells are rarely observed in the heart (Fig 1e) after 1 week of labeling. In subsequent studies, small intestine was used as a positive control to confirm label delivery. To evaluate for an age-related change in cell cycle activity, we administered 15N-thymidine for 8 weeks to three age groups of C57BL6 mice starting at day-4 (neonate), at 10-weeks (young adult) and at 22-months (old adult) (Supplemental Fig 2). We then performed MIMS analysis (Fig 2a, b, Supplemental Fig 3). In the neonatal group, 56% (±3% s.e.m., n=3 mice) of cardiomyocytes demonstrated 15N nuclear labeling, consistent with the well-accepted occurrence of cardiomyocyte DNA synthesis during post-natal development 19 . We observed a marked decline in the frequency of 15N-labeled cardiomyocyte nuclei (15N+CM) in the young adult (neonate= 1.00%15N+CM/day ±0.05 s.e.m. vs young adult=0.015% 15N+CM/day ±0.003 s.e.m., n=3 mice/group, p 320 cardiomyocytes entering the cell cycle; these results exclude such a high rate of turnover (expected 15N+ cardiomyocytes=321; observed=35; Fisher’s exact 2N) in the remaining cardiomyocytes as expected with compensatory hypertrophy after injury. Thus, in the 8wks after myocardial infarction, approximately 3.2% of the cardiomyocytes adjacent to the infarct had unambiguously undergone division (total 15N+ × mononucleated diploid fraction = 23% × 0.14 = 3.2%). The low rate of cardiomyocyte cell cycle completion is further supported by the absence of detectable Aurora B Kinase, a transiently expressed cytokinesis marker, which was detected in rapidly proliferating small intestinal cells but not in cardiomyocytes (Supplemental Fig 10). We also considered the possibility that a subset of 15N+ myocytes that were multinucleated and/or polyploid resulted from division followed by additional rounds of DNA synthesis without division. However, quantitative analysis of the 15N+ population did not identify a subpopulation that had accumulated additional 15N-label as would be expected in such a scenario (Supplemental Fig 11). Together, these data suggest that adult cardiomyocytes retain some capacity to reenter the cell cycle, but that the majority of DNA synthesis after injury occurs in preexisting cardiomyocytes without completion of cell division. If dilution of the GFP+ cardiomyocyte pool cannot be attributed to division and differentiation of endogenous progenitors, do these data exclude a role for progenitors in the adult mammalian heart? These data could be explained by preferential loss of GFP+ cardiomyocytes after injury, a process that we have previously considered but for which we have not found supporting evidence 23 . Such an explanation excludes a role for endogenous progenitors in cardiac repair and would be consistent with data emerging from lower vertebrates 8,26 and the neonatal mouse 27 in which preexisting cardiomyocytes are the cellular source for cardiomyocyte repletion. A second possibility to explain the dilution of the GFP+ cardiomyocyte pool is that injury stimulates progenitor differentiation without division; inevitably, this would lead to exhaustion of the progenitor pool, which if true could explain the limited regenerative potential of the adult mammalian heart. In summary, this study demonstrates birth of cardiomyocytes from preexisting cardiomyocytes at a projected rate of approximately 0.76%/year (15N+ annual rate × mononucleated diploid fraction = 4.4% × 0.17) in the young adult mouse under normal homeostatic conditions, a rate that declines with age but increases by approximately four-fold after myocardial injury in the border region. This study shows that cardiac progenitors do not play a significant role in myocardial homeostasis in mammals and suggests that their role after injury is also limited. Online Methods Mice All experiments were conducted in accordance with the Guide for the Use and Care of Laboratory Animals and approved by the Harvard Medical School Standing Committee on Animals. C57Bl/6 male mice were obtained from Charles River. We generated double transgenic Mer-CreMer-ZEG male mice by crossbreeding cardiomyocyte-specific MerCreMer mice and ZEG mice (Jackson Laboratory). β-galactosidase-GFP is under the control of a cytomegalovirus (CMV) enhancer/chicken actin promoter (Actb); the background strain was C57BL/6J (N7). The background strain of the MerCreMer mice was C57Bl/6SV129. Genotyping was performed by PCR on tail DNA using the following primers: MerCreMer forward: 5′-GTCTGAC TAGGTGTCCTTCT-3′; MerCreMer backward: 5′-CGTCCTCCTGCTGGTA TAG-3′; ZEG forward: 5′-AAGTTCATCTGCACCACCG-3′; ZEG backward: 5′-TCCTTGAAGAAGATGGTGCG-3′; ZEG control forward: 5′-CTAGGCCA CAGAATTGAAAGATCT-3′; and ZEG control backward: 5′-GTAGGTG GAAATTCTAGCATCATCC-3′. To induce Cre recombination and GFP labeling in cardiomyocytes, we injected 4-OH-tamoxifen (provided by a generous gift from Laboratoires Besins), dissolved in peanut oil (Sigma) at a concentration of 5 mg/ml, intraperitoneally into 8-week-old MerCreMer-ZEG mice daily at a dosage of 0.5 mg/day for 14 days. Experimental myocardial infarction Mice were subjected to experimental myocardial infarction as described. Surgeries were performed by a single operator with more than 20 years of experience in the performance of coronary ligation in rodents. In brief, the left coronary artery was permanently ligated approximately 2 mm below the left atrial appendage. For sham operations, the thoracic cavity was opened and the heart exposed, but no intramyocardial sutures were placed. 15N-thymidine (Cambridge Isotopes) was administered at a rate of 20μg/hr via osmotic minipump (Alzet), implanted subcutaneously at the time of experimental myocardial infarction after a single intraperitoneal bolus dose of 500μg. MIMS data acquisition The factory prototype of the NanoSIMS50 as well as a standard NanoSIMS 50 and a large radius NanoSIMS 50L (Cameca, Gennevilliers, France) was used for MIMS analysis as previously described 15 . A focused beam of Cs+ ions was used to sputter a few atomic layers and generate secondary ions from the left ventricular free wall of heart section samples. The Cs+ primary ions were scanned over a raster pattern of either 256 × 256 pixels or 512 × 512 pixels. At each pixel location, the secondary ion intensities for 12C−, 13C−, 12C14N− and 12C15N−were recorded in parallel from the same sputtered volume. The detection of nitrogen requires the use of cluster ions 12C14N− and 12C15N− for 14N and 15N, respectively, due to the low ionization efficiency of nitrogen as N−. MIMS data analysis From a single field image acquisition, we first extracted four image files: the four original quantitative mass images (QMIs; 12C, 13C, 12C14N, 12C15N) and the two ratio images (13C/12C and 12C15N/12C14N), derived from the pixel-by-pixel division of the 13C QMI by the 12C QMI and of the 12C15N QMI by the 12C14N QMI, respectively. We then used a hue saturation intensity transformation (HSI) of the ratio image to map 15N-labeled regions. The hue corresponds to the ratio value, and the intensity at a given hue is an index of statistical reliability. 15N-thymidine labeling For the neonatal cohort in the aging experiment, labeling was starting at post-natal day 4 with subcutaneous injections of 50μg/g 15N-thymidine (Cambridge Isotopes) every 12h and continued through post-natal week 4. Starting at age 4 wks – and in all other experiments using adult mice – osmotic minipumps (Alzet) were implanted subcutaneously, delivering 15N-thymidine (Cambridge Isotopes) at a rate of 20 μg/hour. Multinucleation Analysis Serial adjacent sections (0.5μm) were stained to identify cardiomyocyte borders. A given cardiomyocyte was tracked in the vertical axis by locating it in serial sections. Uninjured hearts were stained using a modified PAS protocol with standard solutions (Electron Microscopy Services), but with longer incubation times optimized for LR white embedding. Slides were incubated in xylene at 37C, 1 hour, rehydrated through graded alcohols, incubated in periodic acid for two hours, and Schiff’s reagent for two nights. Sections were counterstained in hematoxylin and Scott’s Bluing for 1 hour each. Injured hearts were stained using a modified Trichrome staining protocol with standard solutions (Fisher Scientific), but with longer incubation times. Slides were incubated in xylene at 37C, 1 hour, rehydrated through graded alcohols, incubated in bouin’s fluid at 56C, 1 hour, rinsed in tap water, incubated in weigert’s iron hematoxylin stain 1 hour, rinsed in tap water, incubated in scarlet-acid fuchsin solution, 1 hour, rinsed in DI water, incubated in phophotungistic-phophomolybdic acid solution, 30 minutes, incubated in aniline blue stain solution, 30 minutes, and incubated in 1% acetic acid, 20 minutes. Fluorescent in situ hybridization Sections were incubated in proteinase K (50ug/ml) at 60C for 15min. After a PBS/MgCl2 (45mM) wash, slides were post-fixed in 4% paraformaldehyde (PBS/MgCl2), dehydrated through graded alcohols. Biotinylated-labeled chromosome Y paint (Star-FISH, Cambio) in hybridization mix was applied to sections, and sealed under glass with rubber cement (note some samples were analyzed with chromosome-18 paint due to product discontinuation of Y-paint). Samples were heated to 90C for 15 min. After an overnight incubation at 37C, slides were washed three times with 50% formamide/2x standard saline citrate at 45C, three washes with 2x standard saline citrate at room temperature, and two washes with 4x standard saline citrate/0.1% Tween at room temperature. Samples were blocked 10 minutes with 4x standard saline citrate/0.1% Tween/ 0.05% milk and incubated for 2 hr in streptavidin-conjugated Alexa Fluor 488 (Invitrogen) prior to washing and mounting. An observer unaware of the MIMS images or 15N-thymidine labeling status of the nuclei assigned ploidy status. Immunofluorescent staining Sections were incubated in glycine/tris (50mM glycine/0.05M Tris) at room temperature for 5min. After a brief wash with Tris, sections were incubated with chicken anti-GFP (Abcam), rabbit anti-ckit (Abcam), rat anti-sca1 (Abcam) with fresh 0.1%BSA in TBST (TBS/0.1%Tween) overnight at 4C. After a brief wash with TBS, sections were incubated with anti-chicken Alexa Fluor 488 (Invitrogen) prior to TBS wash and mounting. An observer unaware of the MIMS images or 15N-thymidine labeling status of the nuclei assigned GFP status. Fluorescence Image analysis We used a custom-written script in IP Lab version 4.0 (Scanalytics) imaging software for serial image acquisition. Tissue sections were auto-imaged using an Olympus IX-70 microscope with a digital charge-coupled device camera (CoolSNAP EZ, Roper Scientific), an automated stage with a piezoelectric objective positioner (Polytec PI, Auburn MA) to compensate for deviations in the z-axis. Images were compressed and stitched into a mosaic using stitching software (Canon Photostitch). Multichannel images were merged in ImageJ prior to stitching. Statistical analysis Statistical testing was performed using Prism 3.0 (Graphpad). Results are presented as mean ± s.e.m. and were compared using T-tests (significance was assigned for p<0.05). Data comparing event rates were tested with a Fisher-Exact test. Supplementary Material 1