- Record: found
- Abstract: found
- Article: found
Insecticidal decay effects of long-lasting insecticide nets and indoor residual spraying on Anopheles gambiae and Anopheles arabiensis in Western Kenya
Read this article at
Abstract
Background
Indoor residual spraying (IRS) and long-lasting insecticidal nets (LLINs) are the first-line tools for malaria prevention and control in Africa. Vector resistance to insecticides has been extensively studied, however the insecticidal effects of the nets and sprayed walls on pyrethroid resistant mosquitoes has not been studied thoroughly. We evaluated the bioefficacy of LLINs of different ages and lambda-cyhalothrin (ICON 10cs) on the sprayed mud walls for a period of time on malaria vector survivorship.
Methods
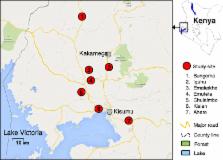
WHO tube bioassay was performed using diagnostic doses of lambda-cyhalothrin (0.05 %), permethrin (0.75 %) and deltamethrin (0.05 %). Cone bioassays were conducted on netting materials from 0 to 3 years old long-lasting insecticide-impregnated nets. Wall bioassays were performed monthly on mud slabs sprayed with lambdacyhalothrin over a period of seven months. All bioassays used An. gambiae mosquitoes collected from the field and the laboratory susceptible reference Kisumu strain. Concentration of the insecticides on the netting materials was examined using the gas chromatography method. Mosquitoes were identified to species level using PCR and genotyped for the kdr gene mutation frequencies.
Results
WHO bioassays results showed that populations from five sites were highly resistant to the pyrethroids (mortalities ranged from 52.5 to 75.3 %), and two sites were moderately resistant to these insecticides (80.4 – 87.2 %). Homozygote kdr mutations of L1014S ranged from 73 to 88 % in An. gambiae s.s. dominant populations whereas L1014S mutation frequencies were relatively low (7–31 %) in An. arabiensis dominant populations. There was a significant decrease ( P < 0.05) in mosquito mortality with time after the spray with both lambda-cyhalothrin (75 % mortality after six months) and with the age of LLINs (60 % mortality after 24 month). Field collected mosquitoes were able to survive exposure to both IRS and LLINs even with newly sprayed walls (86.6–93.5 % mortality) and new LLINs (77.5–85.0 % mortality), Wild mosquitoes collected from the field had significantly lower mortality rates to LLINs (59.6–85.0 %) than laboratory reared susceptible strain (100 %). Insecticide concentration decreased significantly from 0.14 μg/ml in the new nets to 0.077 μg/ml in nets older than 18 months ( P < 0.05).
Conclusion
This study confirms that insecticide decay and developing levels of resistance have a negative contribution to reduced efficacy of ITN and IRS in western Kenya. These factors contribute to decreased efficacy of pyrethroid insectides in ongoing malaria control programs. In order to mitigate against the impact of insecticide resistance and decay it is important to follow the WHO policy to provide the residents with new LLINs every three years of use while maintaining a high level of LLINs coverage and usage. There is also need for urgent development and deployment of non-pyrethroid based vector control tools.
Related collections
Most cited references56
- Record: found
- Abstract: found
- Article: not found
Identification of single specimens of the Anopheles gambiae complex by the polymerase chain reaction.
- Record: found
- Abstract: found
- Article: found
Anopheles gambiae: historical population decline associated with regional distribution of insecticide-treated bed nets in western Nyanza Province, Kenya
- Record: found
- Abstract: found
- Article: not found